7.7 Major Histocompatibility Complex
Role of MHC proteins in the immune response
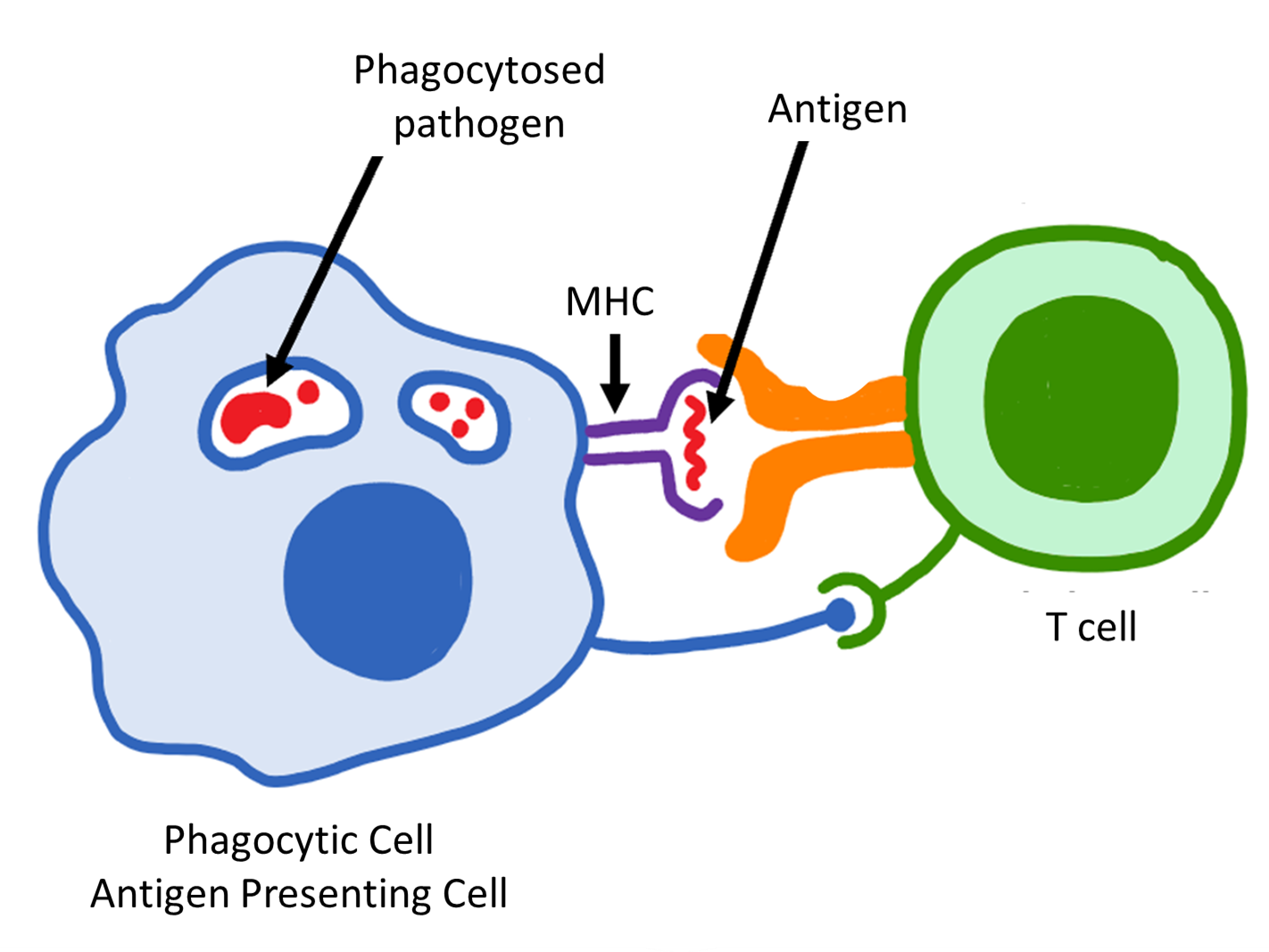
You may be wondering where MHC appears in all of this immune system stuff. We mentioned several times that APCs must present the antigen on their cell surface in order to activate the specific immune response. MHC proteins are the molecules that do the presenting. In other words, MHC proteins bind to the antigen, bring the antigen from inside the cell to face the outside, and then facilitate the binding and activation of T cells (Figure 7.14).
MHC Genetic Diversity
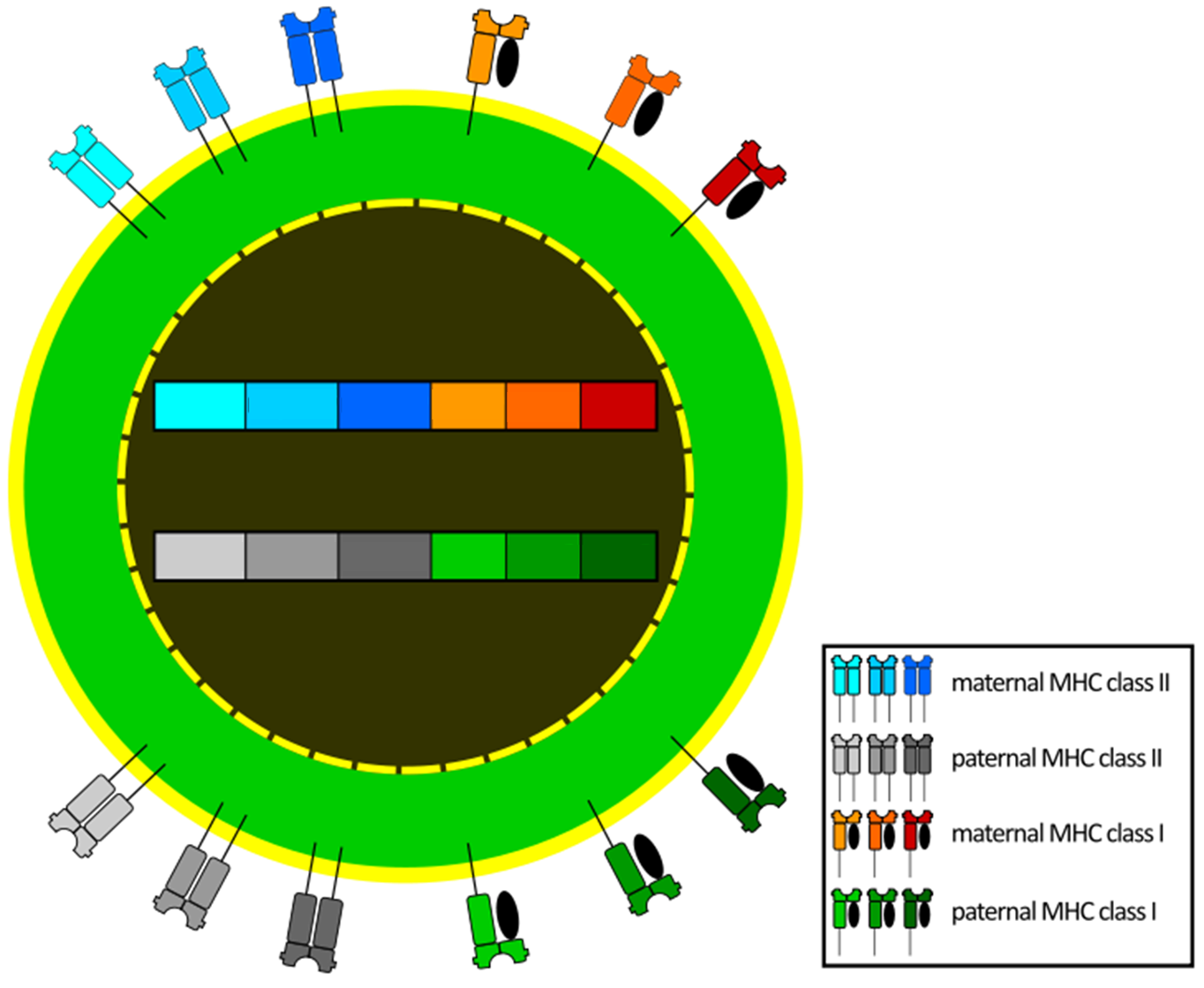
Remember that MHC proteins are part of the bridge between the non-specific and specific immune response. This means that an individual with a lot of variety in their MHC proteins will be able to recognize a broader set of pathogens and therefore be able to effectively fight off the many invaders rather than succumb to the illness.
Genetic diversity of MHC proteins is increased due to the following factors (Figure 7.15):
- MHC proteins involved in antigen presentation are encoded by MHC class I and MHC class II for a total of 6 different MHC genes involved in antigen presentation .
- For each MHC gene there are thousands of alleles.
- MHC gene expression is codominant.
- Most vertebrates are diploid.
Point to Ponder
Consider the ability of an individual that is homozygous for all 6 MHC genes to respond to many kinds of pathogens. Compare that ability to an individual that is heterozygous for all MHC genes. Which one will be more likely to have an effective immune response that clears the invader? Can you explain why?
MHC proteins and Mate Choice
The Red Queen Hypothesis made 3 predictions. Two of them are important to our exploration of the role of MHC proteins in mate choice.
Let’s evaluate the following prediction:
- Pathogens are more likely to attack individuals with common phenotypes (for example, clones) in a population, as opposed to individuals with the less-common phenotypes (such as those that resulted from sex).
How does the genetic diversity of the Major Histocompatibility Complex connect to the above prediction? In your response, make explicit connections to the 4 factors listed above that contribute to the genetic diversity of MHC proteins.
Let’s evaluate the following prediction:
- In sexually reproducing populations, individuals choose mates that maximize diversity in their offspring.
Female sand lizards mate indiscriminately —that is, they seem to be willing to mate with any male who presents himself. One might expect with this type of mating strategy that the paternity of her offspring would be random among the males she has mated with. However, this is not the case. Rather, the male that differs the most from the female at MHC alleles has the highest chance of fathering baby sand lizards.

Some have speculated that however promiscuous the sand lizard is, her reproductive tract is more discriminating. In other words, selection for genetically dissimilar sperm occurs within the female sand lizard reproductive tract. This is an example of “cryptic female choice,” which will be discussed more in Chapter 9.
Even stranger, when a female sand lizard mates with two males, the second male adjusts the amount of sperm-bearing semen he ejaculates. Specifically, the second male appears to evaluate the degree of MHC-relatedness between the previous male and the female. If the first male is similar to the female, the second male has a larger ejaculate than if the first male is less similar to the female. This is an example of “sperm competition,” which will also be discussed more in Chapter 9. This suggests that not only can the female sand lizard detect and discriminate based on MHC similarity, but a sand lizard male can detect the MHC type of the previous mate and adjust his ejaculate accordingly.
But how does he “know”? Key to many studies of MHC-related mate choices is a sense of smell. Apparently, many vertebrates (such as birds, fish, and mammals) can detect MHC complementarity using scent. Sand lizards are no exception; in fact, a characteristic of sand lizard mating is the male’s tendency to sniff the female’s genitalia prior to copulation. This behavior makes sense if the male can sniff out important information about rival males. After all, this information may ultimately increase his fitness.
MHC and Mate Choice in Humans
Do humans have MHC-related mate choices? One study, led by Raphaëlle Chaix, looked at European American spouses and asked whether they were more MHC-dissimilar than random pairs of individuals. [4] They did this by looking at 9,010 different positions within the MHC region of the human genome. Then they measured average relatedness coefficients (r) between spouses.
The coefficient of relatedness (r) is the measure of the probability that two individuals share the same allele due to a recent, common ancestor. In other words, r measures how closely related two individuals are to each other.
Positive values of r indicate genetic similarity and negative values of r indicate genetic dissimilarity, compared to random mating.
The researchers found that the spouses were significantly more MHC-dissimilar from each other than random pairs of individuals (r = -0.043, p = 0.015). This suggests that humans do select for mates based on MHC dissimilarity and therefore increased MHC variation in their offspring.
How would two humans detect MHC similarity? We can’t just ask someone which alleles they have at their MHC loci and compare them to our own. There is some evidence that humans may prefer the smell of a MHC-dissimilar person and that may contribute to mate choice [5]. However, this field is changing and cultural factors in modern humans make this type of study challenging as it is hard to control for cultural differences [6]. There is also some evidence that the cervical mucus in the female’s reproductive tract may select for sperm that are MHC-dissimilar [7].
- Image "Activation of T and B cells" by Immcarle105. Used with CC-BY-SA 4.0 license. ↵
- Image "MHC expression" by Zionlion77. Used with CC-BY-SA 3.0 license. ↵
- Image "Two Lizards" by 2happy, shared with CC-0 license ↵
- 4. Chaix R, Cao C, Donnelly P. Is mate choice in humans MHC-dependent? PLoS Genet. 2008 Sep 12;4(9):e1000184. ↵
- 5. Wedekind, C., Seebeck, T., Bettens, F., & Paepke, A. J. (1995). MHC-Dependent Mate Preferences in Humans. Proceedings of the Royal Society. B, Biological Sciences, 260(1359), 245–249. ↵
- 6. Havlicek, J., Winternitz, J., & Roberts, S. C. (2020). Major histocompatibility complex-associated odour preferences and human mate choice: near and far horizons. Philosophical Transactions of the Royal Society of London. Series B. Biological Sciences, 375(1800), 20190260–20190260. ↵
- 7. Jokiniemi, A., Magris, M., Ritari, J., Kuusipalo, L., Lundgren, T., Partanen, J., & Kekalainen, J. (2020). Post-copulatory genetic matchmaking: HLA-dependent effects of cervical mucus on human sperm function. Proceedings of the Royal Society. B, Biological Sciences, 287(1933), 20201682–20201682. ↵

{kind=link}
{kind=link}